Comparative Morphology

Arkive.org
|

Arkive.org
|
|
Anatomy
The arctic fox is fairly small (3-5 kg) with reduced limb size, a short snout, short and rounded ears, thick haired feet, and a dense winter coat. The tail is bushy and accounts for one third of the total body length (Prestrud 1991). Males tend to weigh 5-20% more than females. There are two colour morphs: blue and white. The blue colour morph is dark throughout the year while the white morph is white in winter and brown dorsally and ventrally throughout the summer. The white morph is more common in foxes living in continental (inland) territories while the blue morph is more common in coastal (marine) habitats (Pagh and Hersteinsson 2008). Dental formula is: i 3/3, c 1/1, p 4/4, m 2/3, for a total of 42, pictured below (Audet et al. 2002). Digestive anatomy: It is similar to that of most carnivorous species, specifically canids. The stomach is simple and the overall length of the small and large intestines is short, and the cecum is small (Stevens and Hume 1995). Image of the digestive system of a dog is pictured below (Stevens and Hume 1998), and is similar to both foxes described here, considering they are closely related (see phylogenetic tree pictured below, Williams et al. 2004). |
Anatomy
The fennec fox is the smallest canid species (with an average weight of 1kg but may weigh as much as 1.5kg). Ears are large (~10cm in length), and muzzle is relatively small. The height at the shoulder is on average 20cm, and the total length rarely exceeds 60cm. The ears are white inside with cinnamon rufous outside, while the central part of the ear is naked internally and covered with short, fine hairs elsewhere. The flanks, underparts, and limbs are almost white. The tail is short (~215 mm), bushy, thick, and often has a black patch near the dorsal base and well as a black tip. The paw pads are completely covered with hairs up to 13mm in length. (Gaulthier-Pilters 1967). Dental formula is: i 3/3, c 1/1, p 4/4, m 2/3, for a total of 42, pictured below (Lariviere 2002). Digestive anatomy: similar to other carnivores and canids (See description in arctic fox Anatomy section). |
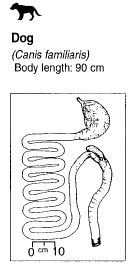
Stevens and Hume 1995
|
Williams et al. 2004
|

Audet et al. 2002
|
Lariviere 2002
|
|
Environmental Adaptations
The major adaptation to the cold climate is undoubtedly the fur. An increase in fur depth of 200% from summer to winter has been reported (Underwood 1971). Parts of the body exposed to the cold have the thickest fur. The legs are not as well insulated as the rest of the body, and its rich blood supply is a potential source of heat loss. This loss of heat is avoided by countercurrent vascular heat exchange, which also prevents freezing of foot pads (Irving and Krog 1955). Calculated by Fuglesteg et al. (2006), the thermal conductivity of fur is between 0.059-0.080 W/m 'C, and the thermal conductance was 2.65 for summer furs and 1.24 for winter furs. Subcutaneous fat may also contribute to insulation, but not nearly to the same degree as the thick fur coat (Prestrud and Nilssen 1992). Short ears and a short muzzle also help to minimize potential heat loss (Macpherson 1969). The arctic fox can also employ behavioural adaptations in response to low ambient temperatures. The fox will curl up and hide its nose under its tail; temperatures up to 26-50'C higher than ambient temperature can be sustained in this position (Follmann 1978). The fox can also dig into snow or seek shelter inside dens. The feet are relatively large compared to body mass, which causes a low foot load that aids locomotion in the snow (Audet et al. 2002). BMR ranges from 35-110 kcal/kg^0.75 (Audet et al. 2002). As calculated BMR is 91.8(4^0.813) = 283 kcal/kg, although due to many adaptations the value found in the literature may be more correct. It is controversial as to whether there is seasonal variation in BMR in the arctic fox (Kleiber 1961). A study by Fuglesteg et al. (2006) found a reduction in BMR from summer to winter, and found that metabolic rate also decreased. This implies a down-regulation of cellular metabolism in winter, which will be adventagous in episodes of starvation. In addition, Fuglesteg et al. (2006) also found that in response to starvation, BMR did not drop. It is likely this occurred because fat stores are able to provide the animals with energy for several weeks during starvation. In fact, arctic foxes with 22% body fat were found to survive at a basal metabolic level for on average 30 days (Prestrud 1992). This storage of fat occurs in the summer and autumn in preparation for food shortages in the winter, and therefore body mass changes seasonally (see image below, Fuglesteg et al. 2006). Temperature: It has been suggested that the lower critical temperature for the arctic fox in winter is -40'C (Scholander et al. 1950), however this is controversial. A study conducted by Fuglesteg et al. (2006) found the average lower critical temperature to be as high as -7'C in winter, and as high as 5'C in summer. The authors found it reasonable to conclude that these values were higher than previously expected, so it should the value of -40'C should not be taken as correct. |
Environmental Adaptations
The major challenge the fennec must face is not the scarcity of water in its habitat, but its intolerance to high ambient temperatures (Noll-Banholzer 1979). Above an ambient temperature of 32'C, their body temperature will rise to a critical point (Noll-Banholzer 1979). Under heat stress, fennecs deprived of water decrease their activity and food intake and consequently will lose weight (Lariviere 2002). Fennec foxes most notable feature is undoubtedly the large ears, which are believed to aid in cooling and locating of prey (Dempsey et al. 2009). Its small size may also assist in desert survival; the "resource limitation hypothesis" states that because smaller animals require less energy, they have an advantage over larger animals in environments with poor resource abundance (Roth 1990). Nocturnal behaviour allows the fennec to avoid the daytime desert heat by sleeping in dens, made capable by their small body size, and be more active when the ambient temperatures are much cooler (Maloiy et al. 1982). Burrows never exceed 34'C and nocturnal temperatures in the Sahara range from 25-30'C (Kirmiz 1962). The soles of the paws are covered by long, soft fur to protect against the hot sand and to facilitate locomotion (Bekoff 1975). Physiological adaptations to reduce water loss and requirements are plenty, and includes a considerably reduced BMR and low minimal conductance, a depressed evaporative water loss, great cardiovascular reserves, and are able to subsist on the body water of their prey alone and produce incredibly concentrated urine (Banholzer 1976). The BMR (0.385 ml/g h, Lariviere 2002, or calculated as 91.8BW^0.813 = 92 kcal/kg) has been measured to be 60.7% of the level predicted by body mass, making it an adaptation to the desert environment as it minimizes endogenous heat load (McNab 1966). Minimal conductance was found to be only 77% of the predicted value based on body mass, which suggests that fennecs are well adapted to low ambient temperatures as well (Noll-Banholzer 1979). Evaporative water loss is low, being only 30% of the predicted value, making it an adaptation to low water availability (Noll-Banholzer 1979). The heart rate of the fennec at thermoneutrality is fairly low and follows a circadian cycle (see image below, Maloiy et al. 1982); it has been recorded to be 98-105 bpm. A low heart rate may support the idea that the capacity of desert animals to maintain a low resting metabolic rate may have to do with characteristics of circulation (Dawson 1973). Temperature: The thermoneutral zone of fennecs ranges from 23-37'C. Panting occurs when temperatures exceed 37'C, and vasodilation occurs (Maloiy et al. 1982). Panting rates of up to 690 breaths/minute have been recorded, and it is likely that this adaptation is used only in emergency situations (Maloiy et al. 1982). |
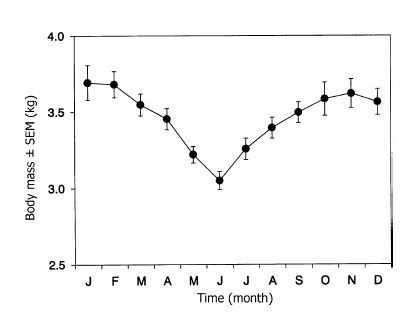
Fuglesteg et al. 2006

Arkive.org
|

Maloiy et al. 1982

Arkive.org
|
